Синтез веществ в почках
В почках образуются некоторые вещества, выделяемые в мочу (например, гиппуровая кислота, аммиак) или поступающие в кровь (ренин, простагландины, глюкоза, синтезируемая в почке, и др.). Гиппуровая кислота образуется в клетках канальцев из бензойной кислоты и гликокола. В опытах на изолированной почке было показано, что при введении в артерию раствора бензойной кислоты и гликокола в моче появляется гиппуровая кислота. В клетках канальцев при дезаминировании аминокислот, главным образом глютамина, из аминогрупп образуется аммиак. Он поступает преимущественно в мочу, частично проникает и через базальную плазматическую мембрану в кровь, и в почечной вене аммиака больше, чем в почечной артерии.
Всасывание воды происходит пассивно, по градиенту осмотического давления и зависит от реабсорбции натрия и хлорида. Реабсорбция натрия в проксимальном отделе осуществляется как активным, так и пассивным транспортом. В начальном участке канальцев это активный процесс
Но небольшое количество изученных испытуемых могло обеспечить недостаточную статистическую мощность, чтобы продемонстрировать увеличение выделения глюкозы в печени. Наши собственные исследования ясно продемонстрировали увеличение выделения глюкозы в печени у пациентов с гипергликемией с диабетом 1 и 2 типа.
Осмотическое разведение и концентрирование мочи
Способностью к осмотическому разведению мочи, т. е. способностью к выделению мочи с меньшей концентрацией осмотически активных веществ, а следовательно, с меньшим осмотическим давлением, чем плазма крови, обладают почки млекопитающих, птиц, рептилий, амфибий, пресноводных рыб и круглоротых. В то же время способностью к образованию мочи с большей концентрацией осмотически активных веществ, т. е. с большей осмотической концентрацией, чем кровь, обладают лишь почки теплокровных животных. Многие исследователи пытались разгадать физиологический механизм этого процесса, но лишь в начале 50-х годов XX века была обоснована гипотеза, согласно которой образование осмотически концентрированной мочи обусловлено деятельностью повороти о-противоточной множительной системы в почке.
Почечная и печеночная высвобождение глюкозы при диабете типа 2. □, недиабетический; ▪ диабетический. Интересно отметить, что оба вышеупомянутых комбинированных исследования баланса изотопных сетей продемонстрировали, что почечная недостаточность глюкозы была увеличена у пациентов с диабетом. Таким образом, некоторые из увеличенного выделения глюкозы в почках у пациентов и животных с диабетом могут быть вызваны увеличением гликогенолиза почек.
Гомеостатическая и метаболическая функции
Недавние исследования показывают, что почка человека потребляет и высвобождает глюкозу в постабсорптивном состоянии и что, следовательно, более нецелесообразно отождествлять выделение глюкозы в печени с общим системным высвобождением глюкозы, измеренным изотопически. Похоже, что почка может быть примерно столь же важна, как и глюконеогенный орган, как печень. Почечная выделение глюкозы и поглощение находятся под гормональным контролем. Почка играет роль в регуляции глюкозы человека, может компенсировать, по крайней мере, частично для ослабленного выделения глюкозы в печени и способствует чрезмерному высвобождению глюкозы, наблюдаемому при обоих типах диабета 1 и 2.
Принцип противоточного обмена достаточно широко распространен в природе и используется в технике. Механизм работы такой системы рассмотрим на примере кровеносных сосудов в конечностях арктических животных. Во избежание больших потерь тепла кровь в параллельно расположенных артериях и венах конечностей течет таким образом, что теплая артериальная кровь согревает охлажденную венозную кровь, движущуюся к сердцу (рис. 12.8, А). В стопу притекает артериальная кровь низкой температуры, что резко уменьшает теплоотдачу. Здесь такая система функционирует только как противоточный обменник; в почке же она обладает множительным эффектом, т. е. увеличением эффекта, достигаемого в каждом из отдельных сегментов системы. Для лучшего понимания ее работы рассмотрим систему, состоящую из трех параллельно расположенных трубок (рис. 12.8, Б). Трубки I и II дугообразно соединены на одном из концов. Стенка, общая для обеих трубок, обладает способностью переносить ионы, но не пропускать воду. Когда в такую систему через вход I наливают раствор концентрации 300 мосмоль/л (рис. 12.8, Б, а) и он не течет, то через некоторое время в результате транспорта ионов в трубке I раствор станет гипотоническим, а в трубке II - гипертоническим. В том случае, когда жидкость течет по трубкам непрерывно, начинается концентрирование осмотически активных веществ (рис. 12.8, Б, б). Перепад их концентраций на каждом уровне трубки вследствие одиночного эффекта транспорта ионов не превышает 200 ммоль/л, однако по длине трубки происходит умножение одиночных эффектов, и система начинает работать как противоточная множительная. Так как по ходу движения жидкости из нее извлекаются не только ионы, но и некоторое количество воды, концентрация раствора все более повышается по мере приближения к изгибу петли. В отличие от трубок I и II в трубке III регулируется проницаемость стенок для воды: когда стенка становится водопроницаемой - начинает пропускать воду, объем жидкости в ней уменьшается. При этом вода идет в сторону большей осмотической концентрации в жидкость возле трубки, а соли остаются внутри трубки. В результате этого растет концентрация ионов в трубке III и уменьшается объем содержащейся в ней жидкости. Концентрация в ней веществ будет зависеть от ряда условий, в том числе от работы противоточной множительной системы трубок I и II. Как будет ясно из последующего изложения, работа почечных канальцев в процессе осмотического концентрирования мочи похожа на описанную модель.
Необходимы дальнейшие исследования для достижения консенсуса относительно вклада почки в общее выделение глюкозы; оценить влияние доступности субстрата, фармакологических агентов и дополнительных гормонов на выделение глюкозы в почках; и оценить роль почек в различных патологических состояниях.
Мы хотели бы поблагодарить сотрудников Центра клинических исследований за отличную техническую помощь и Мэри Литтл за ее превосходную редакционную поддержку. Этот провитамин находится в самых высоких концентрациях в слое базальной и слоистой спинозы кожи и продуцируется в слизистой оболочке кишечника и печени под действием холестериндегидрогеназы.
В зависимости от состояния водного баланса организма почки выделяют гипотоническую (осмотическое разведение) или, напротив, осмотически концентрированную (осмотическое концентрирование) мочу. В процессе осмотического концентрирования мочи в почке принимают участие все отделы канальцев, сосуды мозгового вещества, интерстициальная ткань, которые функционируют как поворотно-противоточная множительная система. Из 100 мл фильтрата, образовавшегося в клубочках, около 60- 70 мл (2/3) реабсорбируется к концу проксимального сегмента. Концентрация осмотически активных веществ в оставшейся в канальцах жидкости такая же, как и в ультрафильтрате плазмы крови, хотя состав жидкости отличается от состава ультрафильтрата вследствие реабсорбции ряда веществ вместе с водой в проксимальном канальце (рис. 12.9). Далее канальцевая жидкость переходит из коркового вещества почки в мозговое, перемещаясь по петле нефрона до вершины мозгового вещества (где каналец изгибается на 180°), переходит в восходящий отдел петли и движется в направлении от мозгового к корковому веществу почки.
Последнее, в свою очередь, может быть синтезировано эндогенно в слизистой оболочке кишечника и печени и поглощено продуктами животного происхождения. Эргостерол первоначально получен из организма растения и поступает в организм человека через потребление растительных продуктов. Сезон Место жительства Воздействие загрязнения воздуха, загрязнение озоном в промышленных условиях На открытом воздухе Использование солнцезащитных кремов с фактором защиты от солнца Телосложение на религиозной почве Цвет кожи и пигментация Кожные заболевания, ожоги Возраст. Поглощение сильно зависит от типа и количества одновременно поставляемых жиров.
Функциональное значение различных отделов петли нефрона неоднозначно. Поступающая из проксимального канальца, в тонкий нисходящий отдел петли нефрона жидкость попадает в зону почки, в интерстициальной ткани которой концентрация осмотически активных веществ выше, чем в корковом веществе почки. Это повышение осмоляльнои концентрации в наружной зоне мозгового вещества обусловлено деятельностью толстого восходящего отдела петли нефрона. Его стенка непроницаема для воды, а клетки транспортируют Cl-, Na+ в интерстициальную ткань. Стенка нисходящего отдела петли проницаема для воды. Вода всасывается из просвета канальца в окружающую интерстициальной ткань по осмотическому градиенту, а осмотически активные вещества остаются в просвете канальца. Концентрация осмотически активных веществ в жидкости, поступающей из восходящего отдела петли в начальные отделы дистантного извитого канальца, составляет уже около 200 мосмоль/кг Н2О, т. е. она ниже, чем в ультрафильтрате. Поступление С1- и Na+ в интерстициальную ткань мозгового вещества увеличивает концентрацию осмотически активных веществ (осмоляльную концентрацию) межклеточной жидкости в этой зоне почки. На такую же величину растет и осмоляльная концентрация жидкости, находящейся в просвете нисходящего отдела петли. Это обусловлено тем, что через водопроницаемую стенку нисходящего отдела петли нефрона в интерстициальную ткань по осмотическому градиенту переходит вода, в то же время осмотически активные вещества остаются в просвете этого канальца.
Транспортировка и распределение в теле
1-альфа-гидроксилаза опосредует вторую стадию гидроксилирования в митохондриях проксимальной почечной канальцы. До сих пор эта реакция гидроксилирования рассматривалась как стадия деградации с образованием неактивных метаболитов.
- Согласно учебной программе по питательной среде Немецкой медицинской ассоциации.
- Физиологические основы, профилактика, терапия.
- В: Современные знания в области питания.
- Для профилактики, консультирования и терапии.
- Ортомолекулярная профилактика и терапия.
Чем дальше от коркового вещества подлиннику почечного сосочка находится жидкость в нисходящем колене петли, тем выше ее осмоляльная концентрация. Таким образом, в каждых соседних участках нисходящего отдела петли имеется лишь небольшое нарастание осмотического давления, но вдоль мозгового вещества почки осмоляльная концентрация жидкости в просвете канальца и в интерстициальной ткани постепенно растет от 300 до 1450 мосмоль/кг НгО.
Не пытаясь найти аргументы в пользу какого-либо взгляда или мнения вообще, аргументы появляются сами по себе, когда мы ограничиваемся наблюдением за тем, что делают органы, которые отвечают за указание на производство и распределение общего блага в теле. Наблюдая за тем, что они делают и как они это делают, вы понимаете, что на самом деле почти все знают, каковы полномочия правящего класса и как они должны их использовать. Это большое облегчение, поскольку оно показывает, что человек может полагаться на свои врожденные критерии и разоблачать рационализацию, с которой он пытается оправдать злоупотребление властью.
На вершине мозгового вещества почки осмоляльная концентрация жидкости в петле нефрона возрастает в несколько раз, а ее объем уменьшается. При дальнейшем движении жидкости по восходящему отделу петли нефрона, особенно в толстом восходящем отделе петли, продолжается реабсорбция С1- и Na+, вода же остается в просвете канальца. В начальные отделы дистального извитого канальца всегда - и при водном диурезе, и при антидиурезе - поступает гипотоническая жидкость, концентрация осмотически активных веществ в которой менее 200 мосмоль/кг Н2О.
В почке образуются вещества депрессорного действия, т.е. снижающие тонус сосудов и артериальное давление - нейтральный депрессорный липид мозгового вещества, простагландины, кинины и др. Их образование получило название "антигипертензивной " функции почек, поскольку ее нарушение может приводить к артериальной гипертензии
Но давайте не будем продвигать события. Таким образом, это проявляется, не нуждаясь ни в чем толковать, как почки удается обусловливать и направлять функционирование тела в целом и как Сердце взаимодействует с ним. Как мы увидим, бывшие медицинские исследователи учли этот факт, когда они изучали физиологические функции водного элемента тысячи лет назад. Из-за его сложности и множественных тонкостей описание функций «Почки» особенно символично и загадочно. Однако, указывая также анатомические структуры, которые принадлежат ему, медицинская литература древнего Китая указывает на физиологические системы, которые соответствуют «почке».
При уменьшении мочеотделения (антидиурезе), вызванном инъекцией АДГ или секрецией АДГ нейрогипофизом при дефиците воды в организме, увеличивается проницаемость стенки конечных частей дистального сегмента (связующий каналец) и собирательных трубок для воды. Из гипотонической жидкости, находящейся в связующем канальце и собирательной трубке коркового вещества почки, вода реабсорбируется по осмотическому градиенту, осмоляльная концентрация жидкости в этом отделе увеличивается до 300 мосмоль/кг Н2О, т. е. становится изоосмотичной крови в системном кровотоке и межклеточной жидкости коркового вещества почки. Концентрирование мочи продолжается в собирательных трубках; они проходят параллельно канальцам петли нефрона через мозговое вещество почки. Как отмечалось выше, в мозговом веществе почки постепенно возрастает осмоляльная концентрация жидкости и из мочи, находящейся в собирательных трубках, реабсорбируется вода; концентрация осмотически активных веществ в жидкости просвета канальца выравнивается с таковой в интерстициальной жидкости на вершине мозгового вещества. В условиях дефицита воды в организме усиливается секреция АДГ, что увеличивает проницаемость стенок конечных частей дистального сегмента и собирательных трубок для воды.
Каковы основные функции почек в организме
И, зная по современной физиологии и функции и механизмы этих систем, разъясняет поэтический язык, используемый китайской медициной для описания функций кондиционирования и направления почек. Согласно китайской медицине, почка - это корень или источник жизни, поскольку она хранит Сущность ее и заботится о ней, «регулируя рождение, рост, развитие и воспроизводство сущности бытия». В случае человечества Сущность Жизни - это генетическое наследство, которое определяет как унаследованные характеристики человеческого рода, так и пола, а также характеристики семейной генетической истории и личностных характеристик.
В отличие от наружной зоны мозгового вещества почки, где повышение осмолярной концентрации основано главным образом на транспорте Na+ и С1-, во внутреннем мозговом веществе почки это повышение обусловлено участием ряда веществ, среди которых важнейшее значение имеет мочевина - для нее стенки проксимального канальца проницаемы. В проксимальном канальце реабсорбируется до 50 % профильтровавшейся мочевины, однако, в начале дистального канальца количество мочевины несколько больше, чем количество мочевины, поступившей с фильтратом. Оказалось, что имеется система внутрипочечного кругооборота мочевины, которая участвует в осмотическом концентрировании мочи. При антидиурезе АДГ увеличивает проницаемость собирательных трубок мозгового вещества почки не только для воды, но и для мочевины. В просвете собирательных трубок вследствие реабсорбции воды повышается концентрация мочевины. Когда проницаемость канальцевой стенки для мочевины увеличивается, она диффундирует в мозговое вещество почки. Мочевина проникает в просвет прямого сосуда и тонкого отдела петли нефрона. Поднимаясь по направлению к корковому веществу почки по прямому сосуду, мочевина непрерывно участвует в противоточном обмене, диффундирует в нисходящий отдел прямого сосуда и нисходящую часть петли нефрона. Постоянное поступление во внутреннее мозговое вещество мочевины, С1- и Na+, реабсорбируемых клетками тонкого восходящего отдела петли нефрона и собирательных трубок, удержание этих веществ благодаря деятельности противоточной системы прямых сосудов и петель нефрона обеспечивают повышение концентрации осмотически активных веществ во внеклеточной жидкости во внутреннем мозговом веществе почки. Вслед за увеличением осмоляльной концентрации окружающей собирательную трубку интерстициальной жидкости возрастает реабсорбция воды из нее и повышается эффективность осморегулирующей функции почки. Эти данные об изменении проницаемости канальцевой стенки для мочевины позволяют понять, почему очищение от мочевины уменьшается при снижении мочеотделения.
То есть, аналогично тому, что в природе элемент воды хранит зародыши Жизни и делает возможным вместе с теплом и солнечным светом прорастание, рост, развитие и размножение различных материальных проявлений из этих микробов, Почка делает в теле. Поскольку почка сообщается со всем организмом через нервную систему и умеет слушать благодаря слуху, он получает необходимую информацию для определения того, что необходимо для определения и направления различных гормональных систем организма, которые участвуют в регуляции почти все его функции, включая обмен веществ, рост, развитие и воспроизводство, гидроэлектрический баланс и поведение.
Прямые сосуды мозгового вещества почки, подобно канальцам петли нефрона, образуют противоточную систему. Благодаря такому расположению прямых сосудов обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания из крови осмотически активных веществ, поскольку при прохождении крови по прямым сосудам наблюдаются такие же изменения ее осмотической концентрации, как и в тонком нисходящем отделе петли нефрона. При движении крови по направлению к вершине мозгового вещества концентрация осмотически активных веществ в ней постепенно возрастает, а во время обратного движения крови к корковому веществу соли и другие вещества, диффундирующие через сосудистую стенку, переходят в интерстициальную ткань. Тем самым сохраняется градиент концентрации осмотически активных веществ внутри почки и прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам определяет количество удаляемых из мозгового вещества солей и мочевины и отток реабсорбируемой воды.
Функции, связанные с заботой о Сущности Жизни, объясняются знанием современной физиологии, что гормональная система, которая направляет активацию различных гормональных систем организма, находится в мозговом «заполнении», образованном шишковидной железой, гипоталамус и гипофиз. И благодаря увеличению наблюдательной способности при использовании всех видов сложных микроскопов мы знаем, как эта директивная система стимулирует другие гормональные системы организма, которые участвуют в регуляции почти всех функций организма.
Директива гормональной системы условий и направляет гормональное производство в других органах через секрецию гормонов, которые стимулируют определенные органические действия. Например, он оказывает влияние на грудь, на яичники, на яички, а также синтезирует гормон, который стимулирует сокращение матки и секрецию молока из груди. Так что, зная это в течение тысяч лет наблюдением и опытом, китайская медицина теперь может объяснить и продемонстрировать, как почка управляет рождением, ростом, развитием и размножением.
В случае водного диуреза функции почек отличаются от описанной ранее картины. Проксимальная реабсорбция не изменяется, в дистальный сегмент нефрона поступает такое же количество жидкости, как и при антидиурезе. Осмоляльность мозгового вещества почки при водном диурезе в три раза меньше, чем на максимуме антидиуреза, а осмотическая концентрация жидкости, поступающей в дистальный сегмент нефрона, такая же - приблизительно 200 мосмоль/кг Н2О. При водном диурезе стенка конечных отделов почечных канальцев остается водопроницаемой, а из протекающей мочи клетки продолжают реабсорбировать Na+. В итоге выделяется гипотоническая моча, концентрация осмотически активных веществ в которой может снижаться до 50 мосмоль/кг Н2О. Проницаемость канальцев для мочевины низкая, поэтому мочевина экскретируется с мочой, не накапливаясь в мозговом веществе почки.
Директивная гормональная система также синтезирует гормоны, которые стимулируют метаболические функции путем синтеза определенных гормонов в щитовидной железе и в надпочечниках. А через гормон роста «почка» стимулирует синтез белков и общий рост почти всех клеток и тканей организма.
Давайте сначала посмотрим, какие гормоны синтезируются в гипоталамусе и гипофизе
Эти гормоны представляют собой молекулы, состоящие из аминокислот, как и все белки, которые могут быть синтезированы любой клеткой. Разница в том, что они имеют тенденцию быть более простыми, поскольку они содержат меньшее количество аминокислот. Несмотря на это, или, скорее, именно из-за этого, гормоны обладают большей способностью к регулированию на большие расстояния, чем объемные белки, которые обладают более специфическими и локальными регуляторными свойствами.
Таким образом, деятельность петли нефрона, конечных частей дистального сегмента и собирательных трубок обеспечивает способность почек вырабатывать большие объемы разведенной (гипотонической) мочи - до 900 мл/ч, а при дефиците воды экскретировать всего 10-12 мл/ч мочи, в 4,5 раза более осмотически концентрированной, чем кровь. Способность почки осмотически концентрировать мочу исключительно развита у некоторых пустынных грызунов, что позволяет им длительное время обходиться без воды.
Как мы видели в статье под названием «Человеческий мир», наш организм имеет всего двадцать аминокислот для синтеза огромного количества различных типов белков, функции которых необходимы для жизни. Они представляют собой очень сложные молекулы, которые придают живому материалу способность к образованию, трансформации, регуляции, защите, регенерации и развитию, а также являются частью основной структуры тканей. Белки могут содержать несколько сотен или тысяч аминокислот и должны обладать их свойствами и структурой для расположения и последовательности аминокислот, из которых они образуются.
Почки (рис. 26-1) выполняют 3 основные группы функций: мочеобразовательную, гомеостатическую и эндокринную.
Мочеобразовательная функция. Почки экскретируют из организма конечные продукты обмена, посторонние вещества и избыточные соединения. Оттекающие ежесуточно от почек 1,5 л вторичной мочи через мочеотводящие пути выводятся из организма (см. главу 27). Именно по отношению к мочеобразовательной функции (точнее, по отношению к вторичной, или дефинитивной моче) применяют термин «экскреция».
И каждая клетка синтезирует эти белки, необходимые для выполнения своих конкретных функций. Аналогично: аминокислоты, вероятно, соответствуют врожденным ценностям нашего вида и важнейшим ценностям, которые мы приобретаем при изучении жизни. И то, что привлекает внимание, заключается в том, что гормоны, синтезируемые директивной гормональной системой, имеют от 3 до максимум 199 аминокислот. Например, только гормон роста и пролактин, который стимулирует развитие молочной железы и секрецию молока, содержат 199 аминокислот.
Проксимальная реабсорбция глюкозы и аминокислот осуществляется с помощью специальных переносчиков
И для стимуляции секреции гипофизарного гормона, стимулирующего синтез гормонов щитовидной железы, гипоталамусу требуется только 3 аминокислоты. Какими будут три человеческие ценности, которые производят в организме - каскадную реакцию, которая влияет на метаболизм почти во всех клетках организма? Связывая аминокислоты с человеческими ценностями, функционирование гормональной системы предлагает глубокие размышления о директивах, которые обычно принимаются в любой человеческой организации.
Конечные продукты обмена: мочевина, мочевая кислота, креатинин, продукты превращений билирубина, порфирины, аммиак, полиамины, гормоны и их метаболиты.
Поддержание гомеостаза (см. главу 28). Почки отвечают за поддержание постоянства состава и объёма жидкостей организма, электролитов и кислотно-щелочного равновесия (КЩР).
Эндокринная функция. Почки синтезируют гормоны, как поступающие в системный кровоток (эритропоэтин, кальцитриол), так и функционирующие локально вазоконстрикторы и вазодилататоры.
Фильтрация, реабсорбция, секреция и внутрипочечный метаболизм
Мочевыделительная и гомеостатическая функции почек - результат 4 сопряжённых и последовательных процессов: фильтрации, канальцевого транспорта (реабсорбция и секреция), а также внутрипочечного метаболизма. Эти базовые процессы развёртываются между кровеносными капиллярами почек и просветом почечных канальцев.
Клубочковая фильтрация (ультрафильтрация, рис. 26-2) происходит в почечных тельцах из просвета капилляров клубочка (первичная капиллярная сеть) в просвет эпителиальной капсулы и приводит к образованию первичной мочи (ультрафильтрат). Каждые сутки обе почки взрослого человека образуют около 180 л первичной мочи.
Рис. 26-1. Мочевыделительная система. Слева: почки, мочеточники, мочевой пузырь, мочеиспускательный канал (уретра). В составе правой почки: 1 - почечная лоханка; 2 - мозговое вещество почки; 3 - корковое вещество почки. Справа: в составе почечного тельца - капиллярный клубочек (кровь в клубочек вливается из приносящей артериолы, оттекает по выносящей артериоле), наружный листок эпителиальной капсулы Боумена-Шумлянского (внутренний листок эпителиальной капсулы представлен подоцитами (не изображены, см. рис. 26-4); кровь из просвета капилляров фильтруется в полость эпителиальной капсулы, фильтрат - первичная моча, см. рис. 26-2), канальцы нефрона и собирательные трубочки - почечные канальцы, по которым от почечного тельца оттекает первичная моча. В канальцах происходят реабсорбция (см. рис. 26-2 и 26-3) и секреция, в результате образуется окончательная (вторичная) моча, поступающая в почечную лоханку.
Канальцевая реабсорбция (рис. 26-2, 26-3) происходит из просвета почечных канальцев в интерстиций и далее в просвет кровеносных капилляров вторичной капиллярной сети (перитубулярные капилляры). Суточный объём реабсорбции около 179 л.
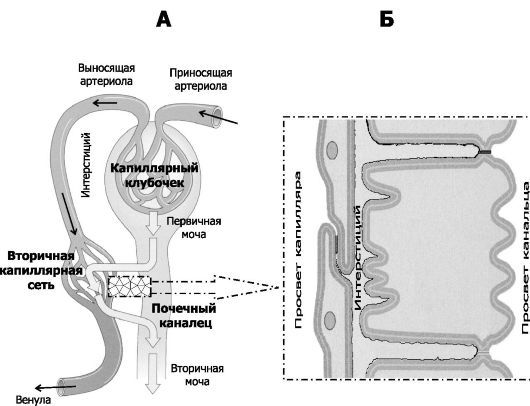 Рис. 26-2. Пути фильтрации, реабсорбции и секреции. А.
Капиллярный
клубочек почечного тельца (первичная капиллярная сеть) перфузируется
артериальной кровью из приносящих артериол. После фильтрации кровь
оттекает от почечного тельца по выносящей артериоле. В интерстиции
(между канальцами) выносящая артериола образует вторичную капиллярную
сеть (перитубулярные капилляры), питающую паренхиму органа. В
перитубулярные капилляры из просвета канальцев происходит реабсорбция, а
из просвета капилляров в просвет канальцев - секреция. В результате из
ультрафильтрата (первичной мочи) образуется дефинитивная моча.
Выделенный прямоугольником участок схематически представлен на Б
Рис. 26-2. Пути фильтрации, реабсорбции и секреции. А.
Капиллярный
клубочек почечного тельца (первичная капиллярная сеть) перфузируется
артериальной кровью из приносящих артериол. После фильтрации кровь
оттекает от почечного тельца по выносящей артериоле. В интерстиции
(между канальцами) выносящая артериола образует вторичную капиллярную
сеть (перитубулярные капилляры), питающую паренхиму органа. В
перитубулярные капилляры из просвета канальцев происходит реабсорбция, а
из просвета капилляров в просвет канальцев - секреция. В результате из
ультрафильтрата (первичной мочи) образуется дефинитивная моча.
Выделенный прямоугольником участок схематически представлен на Б
Канальцевая секреция. Эпителиальные клетки почечных канальцев выделяют в ультрафильтрат ряд химических соединений, поступающих из внеклеточного вещества и перитубулярных капилляров или образующихся в самих эпителиальных клетках канальцев.
Почечный кровоток
Ток крови. Через почечные артерии при каждом сокращении сердца почки получают не менее 20% от сердечного выброса, т.е. около 1200 мл крови в минуту (350 мл/мин на 100 г почечной паренхимы, т.е. почти в 7 раз больше, чем мозг - 50 мл/мин на 100 г ткани мозга).
Почечный ток плазмы крови (именно плазма крови после клубочковой фильтрации образует первичную мочу) составляет примерно 600- 700 мл/мин (при значении Ht - гематокрита - 0,4):
почечный ток плазмы крови = (1 - Нt(почечный кровоток) 600-700 мл/мин = 0,4x1000 мл/мин
Первичная капиллярная сеть. От междольковых артерий параллельно поверхности органа ответвляются короткие приносящие (внутридольковые) артериолы; они распадаются на капилляры, формирующие клубочек в составе почечного тельца - первичная капиллярная сеть (рис. 26-2А и 26-7А). Клубочки первичной капиллярной сети входят в состав почечных телец, в которых происходят фильтрация плазмы и образование клубочкового фильтрата (ультрафильтрата, первичной мочи). Выносящая артериола собирает кровь из капилляров клубочка.
ФОттекающая от почечных телец кровь - артериальная: в выносящей артериоле содержание кислорода лишь примерно на 7% ниже, чем в приносящей артериоле. -Ф- В просвете капилляров первичной капиллярной сети гидростатическое давление составляет примерно 70 мм рт.ст. (вне капилляров, т.е. в полости эпителиальной капсулы - 20 мм рт.ст.), онкотическое - около 30 мм рт.ст. -ФФильтрация в почечных тельцах (рис. 26-2) происходит из просвета капилляров первичной капиллярной сети в полость капсулы Боумена-Шумлянского, движущая сила -
эффективное фильтрационное давление: (гидростатическое давление) - (онкотическое давление) - (давление в полости эпителиальной капсулы) = = (70 мм рт.ст.) - (30 мм рт.ст.) - (20 мм рт.ст.) = 20 мм рт.ст.
Вторичная капиллярная сеть. В капилляры вторичной сети кровь поступает из первичной капиллярной сети через выносящие артериолы. Эти артериолы переходят в прямые артериальные сосуды, спускающиеся в мозговое вещество, образующие вторичную капиллярную сеть (перитубулярные капилляры) и направляющиеся в виде прямых венозных сосудов к корковому веществу. Эти сосуды (и артериальные, и венозные) проходят параллельно канальцам нефронов (канальцы петли Хенле) и собирательным трубочкам, отчего и получили название vasa rectae. Капилляры перитубулярной сети располагаются в непосредственной близости от канальцев нефронов; в эти капилляры реабсорбируются вещества из просвета канальцев (рис. 26-2). Из вторичной капиллярной сети также происходит питание ткани почки. Капилляры мозгового вещества переходят в прямые венулы, впадающие в дуговые вены.
-Φ- Значительное содержание кислорода в капиллярах вторичной капиллярной сети эффективно обеспечивает активную реабсорбцию (рис. 26-2, 26-3) из просвета канальцев в просвет кровеносных капилляров. Кислород необходим главным образом для обеспечения работы Na+,К+-АТФазы, вмонтированной в плазматическую мембрану эпителиальных клеток почечных канальцев. -ФРеабсорбцию поддерживает возросшее в результате фильтрации (по сравнению с капиллярами первичной капиллярной сети) онкотическое давление в капиллярах вторичной капиллярной сети. Итак, первичная капиллярная сеть, расположенная между артериолами, характеризуется высоким гидростатическим внутрикапиллярным давлением и теряет в результате фильтрации не менее 10% объёма крови и до 20% объёма плазмы. Вторичная капиллярная сеть имеет низкое гидростатическое внутрикапиллярное давление, способствующее эффективной реабсорбции из почечных канальцев (см. подробнее на рис. 26- 3Б). Таким образом, вся поступающая в почку артериальная кровь сначала перфузирует капилляры первичной капиллярной сети и лишь затем артериальная кровь поступает в капилляры вторичной капиллярной сети.
Паренхима почки
Паренхима каждой почки, подразделяемая на корковое и мозговое вещество, состоит из 0,8-1,2 млн функциональных структурных единиц - нефронов, а также из множества собирательных трубочек коркового и собирательных протоков мозгового вещества. Коллективно все трубочки почки (канальцы нефрона, собирательные трубочки и протоки) именуются почечными канальцами.
Нефрон - эпителиальная трубка, начинающаяся от почечного тельца и впадающая в собирательную трубку. Стенка нефрона построена из однослойного эпителия, клетки которого (в зависимости от выполняемой функции) различны в разных отделах нефрона. По длине нефрона различают: проксимальный каналец (извитой и прямой) - тонкий каналец петли Хенле - восходящая (толстая) часть петли Хенле (эту часть называют также прямым дистальным канальцем) - извитой дистальный каналец. Дистальный прямой каналец (толстая часть петли Хенле) возвращается к собственному почечному тельцу и контактирует с ним. Извитой дистальный каналец через связующий отдел впадает в собирательную трубочку, которая, в, свою очередь, поступает в собирательные протоки. Разные отделы нефрона закономерно расположены либо в корковом, либо в мозговом веществе.
Типы нефронов. Различают два основных типа нефронов - кортикальные (все отделы нефрона расположены в корковом веществе, 85% всех нефронов - кортикальные) и юкстамедуллярные (петля
Хенле этих нефронов глубоко проникает в мозговое вещество почки).
Отделы почечных канальцев. В нефроне различают несколько отделов: капсула почечного тельца, окружающая капиллярный клубочек; проксимальный извитой и проксимальный прямой канальцы, тонкий каналец (в составе нисходящей и восходящей частей петли Хенле); толстый отдел в составе восходящей части петли Хенле (дистальный прямой каналец), дистальный извитой канальцы, а также связующий отдел (соединяет дистальный отдел нефрона с собирательной трубочкой). Собирательные трубки, сливаясь, образуют собирательные протоки. Характерная особенность всех почечных канальцев состоит в том, что между соседними клетками всегда присутствует диффузионные барьеры в виде полосок плотных контактов, окружающих верхушечные части клеток. Количество таких полосок плотных контактов увеличивается по мере продвижения по почечным канальцам, соответственно увеличивается электрическое сопротивление пласта эпителия, но уменьшается его проницаемость.
Ф- Проксимальный каналец подразделяют на извитой и прямой отделы. Именно в проксимальном отделе нефрона происходит основной объём реабсорбции (рис. 26-3). В связи с этим обстоятельством клетки канальца имеют ряд особенностей, значительно увеличивающим площадь реабсорбции. Интенсивность реабсорбции постепенно уменьшается по мере продвижения первичной мочи по канальцу, соответственно уменьшается количество приспособлений, увеличивающих поверхность клеток, а также митохондрий, необходимых для обеспечения транспортных процессов. По этой причине с функциональной точки зрения (интенсивность реабсорбции) проксимальный каналец подразделяют на последовательные сегменты - S1, S2 и S3. Между соседними клетками встречаются щелевые контакты. Основная функция проксимального канальца - осмос воды, реабсорбция NaCl, NaHCO 3 , глюкозы, аминокислот, Ca 2 +, HPO 4 2- , SO 4 2- , HCO 3 - , а также секреция NH 4 + и некоторых органических катионов и анионов.
Ф- Тонкий каналец петли Хенле состоит из плоских эпителиальных клеток, что существенно уменьшает диффузионный путь для воды. Длина тонкого канальца невелика в кортикальных, но значительна в юкстамедуллярных нефронах. Эти последние (точнее, их петля Хенле), составляя всего 15% от общего количества нефронов, крайне важны для концентрирования или разведения мочи. Клетки петли Хенле перекачивают NaCl из просвета ка-
 Рис. 26-3. Реабсорбция в проксимальном канальце.
Сверху вниз
просвет
канальца, кубические клетки стенки канальца, интерстиций,
перитубулярный капилляр. Стрелки указывают направление и пути
перемещения ионов и молекул: А:
сплошная - трансклеточный
перенос; прерывистая - парацеллюлярный перенос; прерывистые стрелки -
комбинированный вариант переноса (подробнее см. в разделе
«Трансклеточная проницаемость» главы 4, в т.ч. рис. 4-6). 1.
- микроворсинки на поверхности эпителиальной клетки и 2.
глубокие
впячивания в базальной части эпителиальных клеток, а также
переплетающиеся между собой отростки боковых поверхностей соседних
клеток значительно увеличивают поверхность реабсорбции; 3.
Митохондрии
в базальной и латеральной частях эпителиальных клеток необходимы для
обеспечения энергопотребностей при реабсорбции; 4.
плотные
контакты между клетками канальца перекрывают неспецифические пути
диффузии. В верхушечной части клеток в значительном числе находятся
содержащие белок эндоцитозные пузырьки, а также лизосомы. Б.
В
верхней части (эпителиальная клетка) сплошными стрелками показаны пути
трансклеточный, парацеллюлярный и комбинированный пути переноса,
обратите внимание на направленную кверху (в просвет канальца) тонкую
стрелку («утечка» ионов из межклеточного пространства в просвет
канальца). В нижней половине рисунка представлены движущие силы
транспорта через стенку перитубулярного кровеносного капилляра
(параметры: P - гидростатическое давление, π - онкотическое давление;
индексы: i - интерстиций, c - капилляр)
Рис. 26-3. Реабсорбция в проксимальном канальце.
Сверху вниз
просвет
канальца, кубические клетки стенки канальца, интерстиций,
перитубулярный капилляр. Стрелки указывают направление и пути
перемещения ионов и молекул: А:
сплошная - трансклеточный
перенос; прерывистая - парацеллюлярный перенос; прерывистые стрелки -
комбинированный вариант переноса (подробнее см. в разделе
«Трансклеточная проницаемость» главы 4, в т.ч. рис. 4-6). 1.
- микроворсинки на поверхности эпителиальной клетки и 2.
глубокие
впячивания в базальной части эпителиальных клеток, а также
переплетающиеся между собой отростки боковых поверхностей соседних
клеток значительно увеличивают поверхность реабсорбции; 3.
Митохондрии
в базальной и латеральной частях эпителиальных клеток необходимы для
обеспечения энергопотребностей при реабсорбции; 4.
плотные
контакты между клетками канальца перекрывают неспецифические пути
диффузии. В верхушечной части клеток в значительном числе находятся
содержащие белок эндоцитозные пузырьки, а также лизосомы. Б.
В
верхней части (эпителиальная клетка) сплошными стрелками показаны пути
трансклеточный, парацеллюлярный и комбинированный пути переноса,
обратите внимание на направленную кверху (в просвет канальца) тонкую
стрелку («утечка» ионов из межклеточного пространства в просвет
канальца). В нижней половине рисунка представлены движущие силы
транспорта через стенку перитубулярного кровеносного капилляра
(параметры: P - гидростатическое давление, π - онкотическое давление;
индексы: i - интерстиций, c - капилляр)
нальцев в интерстиций, который в результате становится гипертоничным, формируя в мозговом веществе осмотический градиент между корой и почечными сосочками, что имеет решающее значение для осмотической диффузии воды между почечными канальцами и интерстицием.
-Φ- Толстый отдел петли Хенле. Эпителиальные клетки имеют кубическую форму, мощные впячивания плазмолеммы по базальной и латеральной поверхности клеток, что существенно увеличивает поверхность обмена. Это обстоятельство в сочетании с встроенными в плазмолемму клеток характерными трансмембранными переносчиками (см. ниже) существенно важно для формирования гиперосмотической среды. Стенка канальца не проницаема для мочевины и воды.
Ф- Дистальный каналец начинается от плотного пятна (здесь происходит регистрация параметров канальцевой жидкости, подробнее см. ниже) и по своей структуре напоминает клетки толстого отдела петли Хенле.
Ф- Связующий отдел и собирательные трубочки. Их стенка состоит из главных и вставочных клеток. Клетки связующего отдела синтезируют и секретируют калликреин.
♦ Главные клетки несут на свободной поверхности ресничку. Их основная функция - реабсорбция Na + и Cl - и секреция К + .
♦ Вставочные клетки подразделяются на подтипы: A (α) и B (β). Эти клетки реабсорбируют К+. Кроме того, α-клетки секретируют H+, а β-клетки - HCO 3 - .
-Φ- Собирательные протоки. По мере увеличения калибра протоков эпителий становится высоким цилиндрическим, а количество вставочных клеток уменьшается. Собирательные протоки (как и собирательные трубочки) принимают участие в транспорте электролитов, а также под влиянием альдостерона и АДГ - в транспорте воды и мочевины.
Оценка экскреторной функции почек
Для клинической оценки экскреторной функции почек, складывающейся из клубочковой фильтрации, канальцевой реабсорбции и канальцевой секреции, применяют как методы визуализации, так и измерение почечного клиренса (от англ. «clearance» - очищение).
Клиренс
Клиренс вещества X (C X) - параметр, характеризующий выведение почками (экскрецию) из организма вещества X. Клиренс выражают в объёмных единицах за единицу времени (например, в мл/мин). Други-
ми словами, клиренс вещества X - скорость его экскреции, отнесённая к виртуальному объёму крови, полностью очищенной от вещества X.
Для разных веществ значение клиренса (C X) различно. Так, для глюкозы, в норме не экскретируемой, C X равно 0. В то же время для парааминогиппурата, полностью удаляемого из крови, значение C X составляет 700 мл/мин, т.е. равно току плазмы крови через почку.
Клиренс инулина. Некоторые вещества (например, инулин - полимер фруктозы, M r 5000), как и парааминогиппурат, свободно фильтруются, но не реабсорбируются и не секретируются в канальцах. Такие вещества являются хорошим маркёром важного параметра мочевыделительной функции почек - скорости клубочковой фильтрации.
Ф- Скорость клубочковой фильтрации (СКФ, англ. «Glomerular Filtration Rate - GFR») - объём плазмы крови, фильтруемой в единицу времени из крови в полость капсулы Боумена-Шум- лянского (Р х хСКФ).
Для оценки почечного клиренса и СКФ применяют инулин, креатинин, маннитол, 125 1-йоталамат, 57 Co- или 58 Co-цианкобаламин, 51 Cr-этилендиаминтетрауксусную кислоту. Все эти маркёры экзогенны и требуют (в отличие от креатинина) их введения в сосудистое русло обследуемого.
Экскреция
Экскреторную функцию почки по отношению к веществу X (U X xV - скорость экскреции вещества X с мочой) определяют 3 фактора: скорости клубочковой фильтрации (СКФ), канальцевых реабсорбции и секреции. Эти процессы в общем виде можно записать следующим образом: экскреция = фильтрация - реабсорбция + секреция
Экскретируемая фракция (англ. Fractional Excretion - FE) вещества X - полезный показатель оценки функционального состояния почек: отношение скорости экскреции вещества X (U X xV) к объёму клубочковой фильтрации (Рх хСКФ).
ФИЛЬТРАЦИЯ
Через фильтрационный барьер почечного тельца (рис. 26-4, см. также рис. 26-2) происходят фильтрация плазмы и образование первичной мочи (ультрафильтрата, или клубочкового фильтрата).
Фильтрационный барьер
Фильтрационный барьер (рис. 26-4Б,В) состоит из эндотелия капилляров, базальной мембраны и фильтрационных щелей между ножками подоцитов.
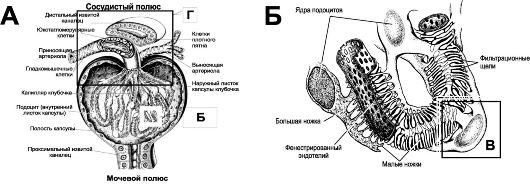 Рис. 26-4. Почечное тельце, фильтрационный барьер и околоклубочковый комплекс
. А. Почечное тельце
состоит
из капиллярного клубочка (примерно 50 капиллярных петель) и
эпителиальной капсулы. Область, где в тельце входит приносящая и выходит
выносящая артериолы, называют сосудистым полюсом; область отхождения
проксимального извитого канальца нефрона - мочевой полюс тельца. Эпителиальная капсула
состоит
из двух листков: наружного (париетального) и внутреннего
(висцерального). Между листками имеется полость, куда из просвета
кровеносных капилляров поступает клубочковый фильтрат. Полость капсулы
открывается в проксимальный извитой каналец. Наружный листок капсулы,
состоящий из однослойного плоского эпителия, ограничивает капсулярное
пространство снаружи. Клетки внутреннего листка капсулы (подоциты)
прикреплены к наружной поверхности капилляров клубочка и вместе с
эндотелием и базальной мембраной, общей для капилляра и подоцитов,
участвуют в процессе фильтрации. К сосудистому полюсу подходит
дистальный извитой каналец того же самого нефрона, что начинается на
мочевом полюсе почечного тельца. Видоизменённые клетки этого отдела
нефрона (плотное пятно) вместе с видоизменёнными клетками приносящей
артериолы (юкстагломерулярные клетки) образуют так называемый околоклубочковый комплекс.
В состав почечного тельца, а также околоклубочкового комплекса также входят мезангиалъные клетки,
расположенные между капиллярными петлями клубочка. Б. Подоциты
-
видоизменённые эпителиальные клетки внутреннего листка капсулы. Они
образуют большие ножки, от которых отходят многочисленные нитевидные
малые ножки. Эндотелиалъные клетки
капилляров клубочка имеют
многочисленные фенестры. Между внутренним листком капсулы и эндотелием
капилляров формируется общая (трёхслойная) базальная мембрана.
Рис. 26-4. Почечное тельце, фильтрационный барьер и околоклубочковый комплекс
. А. Почечное тельце
состоит
из капиллярного клубочка (примерно 50 капиллярных петель) и
эпителиальной капсулы. Область, где в тельце входит приносящая и выходит
выносящая артериолы, называют сосудистым полюсом; область отхождения
проксимального извитого канальца нефрона - мочевой полюс тельца. Эпителиальная капсула
состоит
из двух листков: наружного (париетального) и внутреннего
(висцерального). Между листками имеется полость, куда из просвета
кровеносных капилляров поступает клубочковый фильтрат. Полость капсулы
открывается в проксимальный извитой каналец. Наружный листок капсулы,
состоящий из однослойного плоского эпителия, ограничивает капсулярное
пространство снаружи. Клетки внутреннего листка капсулы (подоциты)
прикреплены к наружной поверхности капилляров клубочка и вместе с
эндотелием и базальной мембраной, общей для капилляра и подоцитов,
участвуют в процессе фильтрации. К сосудистому полюсу подходит
дистальный извитой каналец того же самого нефрона, что начинается на
мочевом полюсе почечного тельца. Видоизменённые клетки этого отдела
нефрона (плотное пятно) вместе с видоизменёнными клетками приносящей
артериолы (юкстагломерулярные клетки) образуют так называемый околоклубочковый комплекс.
В состав почечного тельца, а также околоклубочкового комплекса также входят мезангиалъные клетки,
расположенные между капиллярными петлями клубочка. Б. Подоциты
-
видоизменённые эпителиальные клетки внутреннего листка капсулы. Они
образуют большие ножки, от которых отходят многочисленные нитевидные
малые ножки. Эндотелиалъные клетки
капилляров клубочка имеют
многочисленные фенестры. Между внутренним листком капсулы и эндотелием
капилляров формируется общая (трёхслойная) базальная мембрана.
 Рис. 26-4.
Продолжение.
В. Фильтрационные щели.
Малые
ножки подоцитов прикрепляются к базальной мембране. Между ножками
подоцитов имеются узкие (30-40 нм) фильтрационные щели. Фильтрация
плазмы осуществляется через волокнистую основу базальной мембраны и
фильтрационные щели. Г. Околоклубочковый комплекс
образован тремя
типами клеток, расположенных у корня клубочка. Первый тип -
юкстагломерулярные (зернистые) клетки - видоизменённые и содержащие
гранулы ренина ГМК средней оболочки приносящей артериолы. Второй тип -
юкставаскулярные клетки (мезангиальные), расположенные между приносящей и
выносящей артериолами. Третий тип - эпителиальные клетки дистального
канальца в месте его контакта с корнем клубочка (клетки плотного пятна)
Рис. 26-4.
Продолжение.
В. Фильтрационные щели.
Малые
ножки подоцитов прикрепляются к базальной мембране. Между ножками
подоцитов имеются узкие (30-40 нм) фильтрационные щели. Фильтрация
плазмы осуществляется через волокнистую основу базальной мембраны и
фильтрационные щели. Г. Околоклубочковый комплекс
образован тремя
типами клеток, расположенных у корня клубочка. Первый тип -
юкстагломерулярные (зернистые) клетки - видоизменённые и содержащие
гранулы ренина ГМК средней оболочки приносящей артериолы. Второй тип -
юкставаскулярные клетки (мезангиальные), расположенные между приносящей и
выносящей артериолами. Третий тип - эпителиальные клетки дистального
канальца в месте его контакта с корнем клубочка (клетки плотного пятна)
Эндотелиальные клетки капилляров максимально уплощены, за исключением области, содержащей ядро. Уплощённая часть клетки содержит не затянутые диафрагмой фенестры (овальные окна) полигональной формы диаметром 70 нм, суммарно занимающие примерно 30% всей поверхности эндотелия. В результате плазма крови непосредственно контактирует с базальной мембраной. Таким образом, эндотелиальная часть фильтра задерживает только клеточные элементы, но не плазму крови.
Базальная мембрана толщиной до 300 нм формируется за счёт синтетической активности подоцитов и мезангиальных клеток. Основу базальной мембраны образует мелкоячеистая сеть, образованная молекулами коллагена типа IV, ламинина и связывающих их сульфатированного гликопротеина энтактина. Отрицательно заряженные цепи гепарансульфата, присутствующие в составе протеогликанов базальной мембраны, препятствует прохождению сквозь неё анионов, в том числе и анионных белков плазмы. Вещества с M r до 1 кД проходят через базальную мембрану свободно, до 10 кД в ограниченном количестве, а более 50 кД - в ничтожных количествах.
Фильтрационные щели образованы лабиринтом щелевидных пространств между малыми ножками подоцитов. Фильтрационные щели имеют ширину около 25 нм и затянуты щелевыми диафрагмами (сеть с ячейками размерами от 4 до 14 нм). Щелевые диафрагмы содержат отрицательно заряженные гликопротеины, белок нефрин, а в участках соединения диафрагм с плазмолеммой ножек подоцитов присутствует белок плотных контактов. Ножки подоцитов (за счёт актиновых микрофиламентов) в широких пределах изменяют свою толщину, что неизбежно сказывается на ширине фильтрационных щелей.
Параметры фильтрации
Клубочковую фильтрацию характеризуют различные параметры (объём фильтрата, скорость клубочковой фильтрации - СКФ, эффективное фильтрационное давление, показатель фильтруемости, разности осмотического давления между просветом капилляра и полостью эпителиальной капсулы, характер фильтруемых ионов и молекул).
Объём первичной мочи (отфильтрованной плазмы крови) составляет 10% от объёма крови (20% от объёма плазмы), протекающей по капиллярам клубочка (для взрослого человека 10% от 1800 л крови/сут = 180 л ультрафильтрата/сут, или 125 мл/мин).
СКФ определяют из уравнения:
СКФ = K f xP UF ,
где K f - коэффициент фильтрации; а P UF - эффективное фильтрационное давление.
❖ Коэффициент фильтрации (K f) зависит от гидравлической проводимости клубочковых капилляров и площади фильтрации. При СКФ 125 мл/мин и при P UF 10 мм рт.ст величина K f составляет примерно 12,5 мл/мин/мм рт.ст. (на 100 г массы почки - 4,2 мл/мин/мм рт.ст., что минимально в 200 раз больше, чем K f в любой другой ткани.
♦ Увеличение значения K f повышает СКФ.
♦ Уменьшение значения K f понижает СКФ.
❖ Эффективное фильтрационное давление (P UF , силы Старлинга, или движущая сила фильтрации):
где P GC - гидростатическое давление в просвете клубочковых капилляров (в норме около 50 мм рт.ст. и не изменяется по длине капилляра); P BS - гидростатическое давление в полости капсулы Боумена-Шумлянского (в норме около 10 мм рт.ст.); p GC - онкотическое давление крови в просвете клубочковых капилляров (в начале каждого клубочкового капилляра в норме около 25 мм рт.ст., но постепенно увеличивается, достигая к концу капилляра 30 мм рт.ст.); p BS - онкотическое давление фильтрата в полости капсулы Боумена-Шумлянского (в норме величина этого давления пренебрежимо мала).
Показатель фильтруемости (UF X /P x) - отношение концентрации вещества X в ультрафильтрате (Uf x) к концентрации вещества X в плазме крови - зависит от молекулярной массы и эффективного молекулярного радиуса вещества X.
❖ UF X /P X <1. Вещества с малой молекулярной массой (<5,5 кД) и небольшим эффективным молекулярным радиусом (вода, мочевина, глюкоза, инулин), как правило, имеют в фильтрате ту же концентрацию, что и в плазме крови.
❖ UF X /P X <1. С увеличением молекулярной массы веществ их концентрация в фильтрате прогрессивно уменьшается (например, в ультрафильтрате обнаруживаются лишь следы сывороточного альбумина). Тем не менее, показатель фильтруемости для лизоцима, миоглобина, лактоглобулина и массы других белков с молекулярной массой до 30 кД достаточен для появления в фильтрате ощутимых их количеств.
Электрический заряд. Поскольку ячейки сети в базальной мембране и фильтрационные щели несут отрицательный заряд, это обстоятельство ограничивает фильтрацию анионов и способствует фильтрации катионов. Однако, при этом существенное значение имеют величины молекулярных массы и радиуса заряженных веществ.
Состав клубочкового фильтрата. В результате фильтрации состав первичной мочи оказывается близким к составу плазмы, но в ультрафильтрате нет клеточных элементов крови и относительно мало белка. В частности в первичной моче отсутствуют макромолекулы, эффективный радиус которых превышает 4 нм.
Регуляция почечного кровотока и фильтрации
Имеющие чрезвычайно важное значение для адекватного выполнения функций почек параметры почечного кровотока и фильтрации находятся под жёстким контролем. Известно несколько механизмов контроля кровотока и фильтрации: авторегуляция в виде канальцево-клубочковой обратной связи и эффекты (как сосудосуживающие, так и сосудорасширяющие) множества сосудистоактивных веществ.
Под авторегуляцией понимают не зависящее от нервных и гормональных влияний свойство кровеносной системы почек стабильно удерживать параметры почечного кровотока (следовательно, и СКФ) при значительных колебаниях системного АД (СКФ практически стабильна при систолическом АД 85-150 мм рт.ст.). Авторегуляцию обеспечивают 2 сопряжённых механизма: миогенный ответ ГМК приносящих артериол и канальцево-клубочковая обратная связь.
Миогенный ответ заключается в сокращении или расслаблении ГМК, циркулярно ориентированных по отношению к просвету приносящей артериолы, что приводит к вазоконстрикции или вазодилатации кровеносного сосуда соответственно. Повышение системного АД увеличивает просвет приносящих артериол. Это активирует (открывает) чувствительные к растяжению катионные каналы ГМК, происходит деполяризация плазмолеммы ГМК, поступление Ca 2 + в цитозоль и сокращение ГМК. Просвет сосудов уменьшается, увеличивая сопротивление приносящей артериолы. В результате уменьшается СКФ.
Канальцево-клубочковая обратная связь поддерживается структурами околоклубочкового комплекса.
❖ Околоклубочковый комплекс (рис. 26-4Г) расположен у сосудистого полюса почечного тельца и состоит из юкстагломерулярных клеток, ГМК приносящей артериолы и клеток плотного пятна, принадлежащих стенке дистального извитого канальца того же самого нефрона. Такое тесное соседство ГМК и юкстагломерулярных клеток приносящей артериолы с клетками плотного пятна дистального канальца создаёт хорошие предпосылки для осуществления механизма обратной связи, контролирующего перфузию капиллярного клубочка. В ответ на повышение системного АД возрастает фильтрационное давление и СКФ. При-
рост СКФ увеличивает содержание Na+, Cl - и воды в ультрафильтрате, которое регистрируют клетки плотного пятна и передают соответствующие сигналы к ГМК и юкстагломерулярным клеткам приносящей артериолы.
♦ Клетки плотного пятна реагируют на изменение концентрации и в канальцевой жидкости. Na+/K+/Cl - -перенос- чик, расположенный в плазмолемме верхушечной поверхности клеток плотного пятна, при увеличении в просвете канальца и способствует повышению содержания этих ионов и в цитозоле эпителиальных клеток. В результате открытия катионных каналов плазмолеммы происходит поступление Ca 2 + в цитозоль. Прирост в цитозоле стимулирует секрецию из клеток плотного пятна паракринных и аутокринных агентов в виде аденозина, тромбоксана и некоторых других.
♦ Гладкомышечные клетки стенки приносящей артериолы имеют рецепторы к аденозину, их взаимодействие с выделяющимся из клеток плотного пятна аденозином приводят к поступлению Ca 2+ в цитозоль, сокращению ГМК, вазоконстрикции, увеличению сопротивления приносящей артериолы и уменьшению СКФ.
♦ Зернистые клетки стенки приносящей артериолы также получают сигналы от клеток плотного пятна. Основная функция этих клеток - синтез фермента ренина, поступающего в общий кровоток. Субстрат ренина - ангиотензиноген (см. рис 28-2), дальнейшие превращения которого приводят к появлению в крови ангиотензина II - мощного вазоконстриктора, имеющего и иные эффекты, в том числе и на механизм канальцево-клубочковой обратной связи.
♦ Мезангиальные клетки имеют рецепторы ангиотензина II, атриопептина и вазопрессина. Вазопрессин и ангиотензин II стимулируют сокращение мезангиальных клеток. Так как в цитоплазме клеток в большом количестве присутствуют микрофиламенты, то клетки обладают сократительной активностью и способны уменьшать площадь наружной поверхности стенки капилляров, через которую происходит фильтрация, снижая таким образом её уровень.
Сосудистоактивные регуляторы. В регуляции почечного кровотока и СКФ принимает участие множество гормонов и нейромедиаторов: ангиотензин II, норадреналин, адреналин, дофамин, АДГ, атриопептин, эндотелины, Пг, лейкотриены и оксид азота.
ТРАНСПОРТ В ПОЧЕЧНЫХ КАНАЛЬЦАХ
Транспорт через эпителиальные трубочки в общем виде рассмотрен в разделе «Трансклеточная проницаемость» главы 4 и проиллюстрирован на рис. 4-6 и 26-3. В данном разделе описано канальцевое (трансэпителиальное) перемещение конкретных веществ, т.е. их реабсорбция (из просвета канальцев в интерстиций и далее в околоканальцевые кровеносные капилляры) и секреция (из просвета капилляров в интерстиций и далее в просвет канальцев).
Натрий
Из поступающих в организм при сбалансированной диете 120 ммоль Na+ лишь 15% удаляется через потовые железы и ЖКТ, а 85% экскретируется с мочой. Каждые сутки почки отфильтровывают 25 500 ммоля и реабсорбируют 25 400 ммоля Na + , что примерно эквивалентно полутора килограммам поваренной соли. Поскольку вода пассивно перемещается между компартментами вслед за Na+ (и сопутствующим Cl -), ясно, сколь большое значение имеют почки для поддержания объёма жидкостей организма и их осмоляльности.
Градиент реабсорбции (рис. 26-5А). Реабсорбция Na+ (также CI -) наибольшая в проксимальном извитом канальце, постепенно уменьшается в проксимо-дистальном направлении и наименьшая в собирательных протоках.
Пути и направление транспорта. Через стенку почечного канальца транспорт Na + и Cl - (как и транспорт других ионов и воды) происходит как трансклеточно (сквозь клетку), так и по околоклеточному (парацеллюлярному) пути (рис. 26-3 и 26-5Б,В).
Механизмы канальцевого переноса Na+ через верхушечную и базолатеральную плазмолемму различны в силу того обстоятельства, что существенно отличается внеклеточная (внутриканальцевая и интерстициальная) и внутриклеточная . При реабсорбции Na+ пассивно входит в цитозоль через верхушечную плазмолемму, так как внутриклеточная существенно ниже внутриканальцевой (15 мМ против 142-40 мМ в разных отделах канальцев, см. рис. 26-9А,Б). В то же время через базолатеральную клеточную мембрану Na+ активно выкачивается из клетки, так как внеклеточная, т.е. интерстиция существенно выше внутриклеточной (145 мМ против 15 мМ, см. рис. 26-5Б).
Регуляция канальцевого транспорта Na+ осуществляется по следующим направлениям: автоматическая коррекция реабсорбции в проксимальных и дистальных канальцах вследствие изменений почечного кровотока (следовательно, фильтрации) и влияние на реабсорбцию в дистальном и отчасти в проксимальном отделах нефрона, а также в петле Хенле и особенно в собирательных трубках
 Рис. 26-5. Реабсорбция натрия
. А. Реабсорбция Na+ в разных отделах почечных канальцев.
Стрелки
в просвете канальцев - направление движения фильтрата. Концентрация Na+
в просвете проксимального извитого канальца, петли Хенле, дистального
извитого канальца нефрона и в собирательных трубочках и протоках
приведена в миллимолях (мМ, в прямых скобках). Количество
реабсорбируемого Na+ в сутки (миллимоль/сут и % от отфильтрованного Na+)
приведено внутри стрелок, направленных из просвета канальца. Указана
также трансэпителиальная разность электрохимического потенциала (Δμ Νί1 , мВ).
Рис. 26-5. Реабсорбция натрия
. А. Реабсорбция Na+ в разных отделах почечных канальцев.
Стрелки
в просвете канальцев - направление движения фильтрата. Концентрация Na+
в просвете проксимального извитого канальца, петли Хенле, дистального
извитого канальца нефрона и в собирательных трубочках и протоках
приведена в миллимолях (мМ, в прямых скобках). Количество
реабсорбируемого Na+ в сутки (миллимоль/сут и % от отфильтрованного Na+)
приведено внутри стрелок, направленных из просвета канальца. Указана
также трансэпителиальная разность электрохимического потенциала (Δμ Νί1 , мВ).
 Рис. 26-5.
Продолжение.
Б. Механизм транспорта Na+ в проксимальном извитом канальце (сегмент S1).
Реабсорбция
Na+ из просвета канальца в интерстиций показана стрелками,
направленными слева направо (трансклеточный путь). Na+ входит в клетку
по его концентрационному градиенту (приведены значения концентрация Na+ в
миллимолях), но выходит из клетки в интерстиций против градиента
концентрации. В нижней части рисунка показано обратное движение Na+ по
парацеллюлярному пути, частично замкнутому плотными контактами (показаны
в верхней части рисунка). В верхней части рисунка дана электрическая
схема зарядов в разных частях канальца, из которой следует неизбежность
движения катионов в просвет канальца по парацеллюлярному пути. В. Механизм реабсорбции Na+ в толстом отделе петли Хенле.
Реабсорбция Na+ из просвета канальца в интерстиций показана стрелками, направленными слева направо (трансклеточный путь). Na +
входит в клетку по его концентрационному градиенту, но выходит из
клетки в интерстиций против градиента концентрации. В нижней части
рисунка показана реабсорбция Na + по парацеллюлярному пути,
частично замкнутому плотными контактами (показаны в верхней части
рисунка). В верхней части рисунка дана электрическая схема
трансэпителиальной стенки (сравни с рис. 26-5Б), из которой следует
неизбежность движения катионов из просвета канальца по парацеллюлярному
пути. Положительный заряд на канальцевой поверхности эпителия зависит от
работы множества калиевых каналов, по которым из цитозоля в просвет
канальца поступает K+
Рис. 26-5.
Продолжение.
Б. Механизм транспорта Na+ в проксимальном извитом канальце (сегмент S1).
Реабсорбция
Na+ из просвета канальца в интерстиций показана стрелками,
направленными слева направо (трансклеточный путь). Na+ входит в клетку
по его концентрационному градиенту (приведены значения концентрация Na+ в
миллимолях), но выходит из клетки в интерстиций против градиента
концентрации. В нижней части рисунка показано обратное движение Na+ по
парацеллюлярному пути, частично замкнутому плотными контактами (показаны
в верхней части рисунка). В верхней части рисунка дана электрическая
схема зарядов в разных частях канальца, из которой следует неизбежность
движения катионов в просвет канальца по парацеллюлярному пути. В. Механизм реабсорбции Na+ в толстом отделе петли Хенле.
Реабсорбция Na+ из просвета канальца в интерстиций показана стрелками, направленными слева направо (трансклеточный путь). Na +
входит в клетку по его концентрационному градиенту, но выходит из
клетки в интерстиций против градиента концентрации. В нижней части
рисунка показана реабсорбция Na + по парацеллюлярному пути,
частично замкнутому плотными контактами (показаны в верхней части
рисунка). В верхней части рисунка дана электрическая схема
трансэпителиальной стенки (сравни с рис. 26-5Б), из которой следует
неизбежность движения катионов из просвета канальца по парацеллюлярному
пути. Положительный заряд на канальцевой поверхности эпителия зависит от
работы множества калиевых каналов, по которым из цитозоля в просвет
канальца поступает K+
и протоках гуморальных и нервных факторов, увеличивающих или уменьшающих канальцевый транспорт Na+.
-Φ- Факторы, увеличивающие реабсорбцию Na+, т.е. приводящие к задержке Na+ и воды в организме: альдостерон, АДГ и влияния симпатического отдела нервной системы.
Ф- Факторы уменьшающие реабсорбцию Na+, т.е. приводящие к усилению диуреза и потенциально могущие привести к потере Na+ и обезвоживанию организма: атриопептин, Пг, брадикинин, дофамин и эндогенный ингибитор Na+,К+-АТФазы.
Хлор. Реабсорбция Cl - происходит как по трансклеточному, так и по околоклеточному пути. Объёмы реабсорбции Cl - в разных отделах почечных канальцев практически такие же, как и для Na+ (см. рис. 26-5Б). Cl - поступает в цитозоль против концентрационного градиента путём обмена внеклеточного Cl - на внутриклеточные анионы. Выход Cl - в интерстиций по всему протяжению почечных канальцев обеспечивают Cl - -каналы, а в проксимальном отделе нефрона дополнительно K+/Cl - -контранспортёр.
Вода. Реабсорбция воды по всему протяжению почечных канальцев происходит только пассивно. Из 170 л отфильтрованной воды в проксимальных канальцах реабсорбируется 67%, в петле Хенле - 15%, от 10 до 15% - в собирательных трубках и протоках, не происходит реабсорбции воды в дистальном канальце нефрона. Реабсорбцию воды обеспечивают мембранные водные поры - аквапорины разных типов. Различные ЛС (диуретики), подавляя реабсорбцию Na+, увеличивают экскрецию и Na+, и воды, тем самым уменьшая в организме объём внеклеточной жидкости.
Калий. Почки ежесуточно отфильтровывают 800 мМ К+, хотя с пищей поступает около 100 мМ, а экскретируется с мочой примерно 90 мМ. Происходит также секреция К+. Таким образом, поддержание калиевого баланса организма происходит при сочетании фильтрации, реабсорбции и секреции. В проксимальном отделе нефрона происходит массовая реабсорбция К+ (80%), а в дистальных - в зависимости от поступления калия в организм - этот катион либо реабсорбируется, либо экскретируется. Увеличивают секрецию калия диуретики, низкая в просвете канальцев, альдостерон.
Мочевина. Мочевина - конечный продукт катаболизма аминокислот - образуется в печени из NH 4 +, её концентрация в крови (азот мочевины) - 2,5-8,32 ммоль/л. Все 100% мочевины фильтруется в почках, экскретируется с мочой около 40% отфильтрованной мочевины (ежесуточно 20-35 г). В почках мочевина и реабсорбируется (проксимальный отдел нефрона и собирательные протоки), и секретируется (тонкая часть петли Хенле), в итоге почку покидает венозная кровь, содержащая 5% от поступившей в почки мочевины (рис. 26-6).
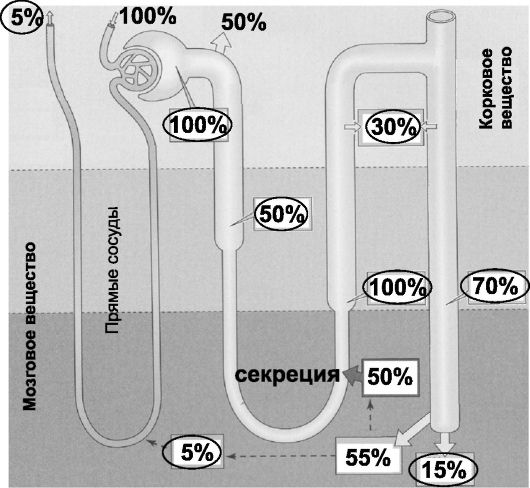 Рис. 26-6. Транспорт мочевины между канальцами, интерстицием и кровеносными сосудами
. Значения в овалах - содержание (в %) от отфильтрованного (100%)
Рис. 26-6. Транспорт мочевины между канальцами, интерстицием и кровеносными сосудами
. Значения в овалах - содержание (в %) от отфильтрованного (100%)
Глюкоза. Концентрация глюкозы в плазме крови натощак - 4-5,5 мМ (3,58-6,05 ммоль/л, 85-115 мг%). В почках глюкоза отфильтровывается полностью и практически полностью и активно (против концентрационного градиента) реабсорбируется в начальных отделах проксимального отдела нефронов. Секреции глюкозы нет, поэтому с мочой экскретируются следовые количества этого сахара. Глюкоза поступает в эпителий канальцев посредством активного сочетанного транспорта с Na+ (электрогенные контранспортёры SGLT), а покидает клетки облегчённой диффузией через Na+-независимые транспортёры GLUT.
Аминокислоты. Концентрация L-аминокислот в крови около 2,4 мМ. Это преимущественно всосавшиеся в ЖКТ аминокислоты. В почках отфильтровываются все аминокислоты, 98% - всасывается в проксимальных извитых канальцах по трансклеточному пути при помощи различныхNa+-зависимых котранспортёров и Na+-независимой облег-
чённой диффузии, выход аминокислот в межклеточное пространство происходит по механизму облегчённой диффузии.
Олигопептиды и белки
Фильтрация. Считают, что макромолекулы с мол. массой выше 40 000 не отфильтровываются. Однако, этот ориентировочный порог не абсолютен. Например, концентрация альбуминов в фильтрате очень низка (от 4 до 20 мг/л, т.е. от 0,01% до 0,05% от концентрации альбумина в плазме крови); тем не менее, при СКФ 180 л/сут, количество отфильтрованного альбумина составляет 0,7-3,6 г/сут. В то же время экскреция альбумина с мочой - около 30 мг/сут. Таким образом, реабсорбируется до 99% отфильтрованного альбумина.
Реабсорбция. Перенос олигопептидов через щёточную каёмку осуществляют Н+-зависимые котранспортёры, тогда как белки поступают в клетки путём опосредованного рецепторами эндоцитоза. Эндоцитозные пузырьки сливаются с лизосомами, где происходит гидролиз белков до аминокислот и олигопептидов. Олигопептиды расщепляются пептидазами до аминокислот как в щёточной каём- ке, так и в цитоплазме эпителиальных клеток. Аминокислоты поступают в интерстиций по механизму облегчённой диффузии. Карбоновые кислоты. Монокарбоксилаты (лактат, пируват, ацетоа-
цетат, β-гидроксибутират), соли ди- и трикарбоновых кислот (α-кетоглутарат, малат, сукцинат и цитрат) реабсорбируются трансклеточно практически полностью в проксимальных извитых канальцах. Экскреция карбоновых кислот - кетоновых тел (ацетоацетат и β-гидроксибутират - происходит при голодании и сахарном диабете.
Органические анионы. Различные органические анионы (метаболиты эндогенно катаболизируемых соединений и экзогенно поступивших ЛС, а также парааминогиппуровая кислота) как фильтруются, так и секретируются. Секреция этих анионов (в том числе оксалатов, солей жёлчных кислот, пенициллина) происходит в проксимальных и дистальных отделах нефрона при помощи анионообменников (в обмен на Cl - , ураты и OH - просвета канальцев).
Ураты - моновалентные анионы - конечный продукт катаболизма пуринов. Их концентрация в плазме крови - 3 -7 мг% (0,2-0,4 мМ). Почки отфильтровывают ураты, в проксимальном отделе нефронов реабсорбируют их (пассивная диффузия и активный транспорт), затем снова происходит их секреция и повторная реабсорбция. С мочой экскретируется примерно 10% отфильтрованных уратов.
Органические катионы (как множество эндогенных (в том числе нейромедиаторы и креатинин), так и экзогенных (например, морфин,
хинин, амилорид) секретируются на протяжении второй половины проксимального отдела нефрона. Их поглощение из интерстиция происходит при помощи облегчённой диффузии, а выход в просвет канальцев осуществляет протонно-катаонный обменник.
Фосфаты. Концентрация фосфатов в плазме крови - 4,2 мг%, 50% находится в ионизированной форме (HPO 4 2- - четыре пятых, H 2 PO 4 - - одна пятая), 40% - в электролитных комплексах, 10% связаны с белками. В почках фильтруются фосфаты в ионизированной и комплексной формах. Ежесуточно фильтруется примерно на порядок величины больше содержания фосфатов в межклеточной жидкости и почти столько же реабсорбируется в проксимальном отделе нефрона при помощи котранспортёра натрия и фосфатов. Гормон паращитовидной железы ингибирует активность этого транспортёра. Некоторое количество фосфатов секретируется в просвет канальцев.
Кальций. Концентрация элементного кальция в плазме крови - 2,2-2,7 мМ. Около 40% кальция связано с белками и в почках не фильтруется, 60% кальция фильтруется из крови, это кальций карбонатов, цитратов, фосфатов и сульфатов (15%) и ионизированный кальций (45%, 1,0-1,3 мМ). 99,5% отфильтрованного кальция реабсорбируется: 65% в проксимальном отделе (этот процесс происходит автоматически и гормонально не контролируется), 35% в толстом отделе петли Хенле и дистальных извитых канальцах (в этих канальцах происходит гормональный контроль реабсорбции Ca 2 +). Гормон паращитовидной железы и витамин D стимулируют реабсорбцию Ca 2 +, тогда как в плазме крови - подавляют реабсорбцию Ca 2 +.
Магний. Концентрация магния в плазме крови - 0,8-1,0 мМ (1,8- 2,2 мг%), 30% магния связано с белками. 70% магния фильтруется в почках: из них менее 10% находится в составе фосфатов, цитратов и оксалатов, 60% - ионизированный магний (Mg 2 +). Менее 5% отфильтрованного магния экскретируется с мочой, 95% реабсорбируется преимущественно по околоклеточным путям во всех отделах нефрона, но главным образом (70%) в толстом восходящем колене петли Хенле. Гормон паращитовидной железы усиливает реабсорбцию во всех канальцах нефрона.
КОНЦЕНТРИРОВАНИЕ И РАЗВЕДЕНИЕ МОЧИ
Почки могут выделять мочу в широком диапазоне осмоляльности: от разведённой (до 30 мОсм, 1/10 осмоляльности плазмы крови) и до концентрированной (до 1200 мОсм, в 4 раза более осмоляльности плазмы). Концентрирование и разведение мочи существенно зависит от баланса воды в организме, транспорта воды, натрия и мочевины в паренхиме почек и специфической организации прямых трубочек (пря-
мых сосудов и петли Хенле) в мозговой части почек в сочетании с избирательной проницаемостью разных отделов петли Хенле и дистальных почечных канальцев.
Баланс воды. В норме поступление воды в организм и потери воды организмом должны быть одинаковы. Поступление воды складывается из выпиваемой жидкости, содержащейся в пище воды и воды, образующейся в митохондриях при аэробном обмене. Потери воды происходят преимущественно через почки, и именно почки - главный регулятор водного обмена.
Постоянство экскреции электролитов. Почки регулируют количество экскретируемой воды в зависимости от экскретируемых электролитов, главным образом хлористого натрия. Константа и стандарт экскреции электролитов - 600 миллиосмоль в сутки. Нормально эти 600 мОсм выделяются с обычными 1500 мл мочи. Для выделения большего или меньшего количества воды почки должны продуцировать мочу иной осмоляльности, но при условии сохранения количества экскретируемых электролитов (600 мОсм). Например, для суточной секреции 600 мОсм в 1500 мл осмоляльность мочи (изоосмотическая моча) должна составлять 400 мОсм; для выделения избытка воды осмоляльность мочи (разведённая моча) может уменьшиться до 30 мОсм (тогда диурез составит 20 л); для сохранения воды почки могут увеличить осмоляльность мочи до 1200 мОсм (диурез - 0,5 л). Таким образом, почки могут развести мочу (по отношению к осмоляльности плазмы крови) примерно в 10 раз (300 и 30 мОсм), но концентрируют мочу только в 4 раза (300 и 1200 мОсм).
Осмоляльность фильтрата в почечных канальцах (рис. 26-7).
Ф- Концентрированная моча образуется при осмотическом перемещении воды из просвета канальцев через водопроницаемые сегменты канальцев в гиперосмотический интерстиций.
Ф- Разведённая моча образуется при транспорте электролитов из просвета канальцев через непроницаемые для воды сегменты.
Ф- Осмотичность фильтрата. В проксимальных отделах нефрона жидкость в просвете канальцев изоосмотична (300 мОсм), после прохождения по петле Хенле - гипоосмотична (120 мОсм), и (в зависимости от баланса воды в организме) в конце собирательных протоков либо гипоосмотична (60 мОсм, рис. 26-7Б), либо гиперосмотична (1200 мОсм, рис. 26-7А).
Гиперосмотичность интерстиция мозгового вещества. Из рис. 26-7 видно, что (как при образовании гипоосмотичной, так и гиперосмотичной мочи) осмоляльность интерстиция мозгового вещества почки всегда выше осмоляльности коркового вещества. Более того,
 Рис. 26-7. Осмоляльность интерстициальной жидкости вокруг разных отделов почечных канальцев
. А.
При ограничении питья. Б.
Обильное питьё
Рис. 26-7. Осмоляльность интерстициальной жидкости вокруг разных отделов почечных канальцев
. А.
При ограничении питья. Б.
Обильное питьё
существует градиент увеличения осмотичности интерстиция в направлении от коркового к мозговому веществу.
Петля Хенле юкстамедуллярных нефронов играет ключевую роль в разведении и концентрировании мочи. Одна из функций петли Хенле состоит в перемещении NaCl из канальцев в интерстиций. В то же время толстый отдел петли не реабсорбирует воду. Тем самым этот сегмент нефрона прямо участвует в образовании разведённой мочи. В то же время возникающая гиперосмотичность интерстиция мозговой части почки косвенно способствует образованию концентрированной мочи (сравни А и Б на рис. 26-7). Такое перемещение NaCl в интерстиций при одновременной непроницаемости для воды толстого отдела петли Хенле в любой точке петли создаёт поперечный градиент осмоляльности между канальцем и интерстицием, равный 200 мОсм. Этого градиента явно мало для создания реально возникающей в месте перегиба петли осмоляльности от 500 мОсм (рис. 26-12Б) до 1200 мОсм (рис. 26-7А). Но эта задача решается повторением циклов создания поперечного градиента между просветом канальца и интерстицием (противоточный умножитель).
Противоточный умножитель петли Хенле. Умножение эффекта создания поперечного градиента осмоляльности возможно в ситуации противоположного движения жидкости в нисходящем и восходящем коленах петли Хенле. Так, осмоляльность в 1200 мОсм в просвете канальца в месте перегиба петли может быть достигнута при повторении цикла более 30 раз. Соответственно нарастает и вертикальный (от коркового к мозговому веществу) градиент осмоляльности (рис. 26-7А). Таким образом, чем длиннее петля Хенле, тем больше вертикальный градиент осмоляльности. Помимо транспорта NaCl в интерстиций из канальцев существенное значение для гиперосмотичности интерстиция имеют и особенности распределения мочевины в разных отделах почечных канальцев (см. рис. 26-6).
Роль прямых сосудов. Прямые сосуды мозгового вещества, расположенные параллельно канальцам петли Хенле и организованные, как и петля Хенле, по типу шпильки (нисходящий прямой сосуд спускается в мозговое вещество, а восходящий прямой сосуд поднимается после перегиба к корковому веществу), также важны для формирования вертикального градиента гиперосмоляльности мозгового вещества. На рис. 23-8 приведены модели противоточного обмена водой и NaCl между просветом сосудов и интерстицием: схема на рис. 26-8А отражает ситуацию только для одного сосуда, а на рис. 26-8Б - для реальной шпильки из нисходящего и восходящего сосудов. Значение прямых сосудов в концентрировании и разведении мочи, как и петли Хенле, состоит в поддержании возрастающе-
го от коры к почечным сосочкам вертикального градиента гиперосмоляльности интерстиция. Здесь имеют значение 2 момента: вопервых, наличие противоточного обмена (сравни с противоточным умножителем градиента в петле Хенле), во-вторых, относительно низкий кровоток в мозговом сравнительно с корковым веществом - не более 10% от объёма кровотока почки. Понятно, что чем ниже кровоток, тем меньше будет удалено электролитов из интерстиция и тем стабильнее будет гиперосмоляльный градиент в мозговом веществе.
 Рис. 26-8. Модель противоточного обмена
. А.
Вертикальная прямая трубка. Б.
Вертикальная
шпилька (петля). Числовые значения - осмоляльное давление (мОсм),
толстые стрелки - движение воды, тонкие стрелки - движение электролитов
Рис. 26-8. Модель противоточного обмена
. А.
Вертикальная прямая трубка. Б.
Вертикальная
шпилька (петля). Числовые значения - осмоляльное давление (мОсм),
толстые стрелки - движение воды, тонкие стрелки - движение электролитов
Собирательные протоки мозгового вещества также важны для формирования гиперосмотичной или гипоосмотичной мочи (рис. 26-7), так как их проницаемость имеет регулируемый характер. Так, без стимулирующего влияния АДГ стенка протоков относительно непроницаема для воды, АДГ увеличивает проницаемость (т.е. реабсорбцию) стенки протоков для воды. И, наконец, АДГ увеличивает проницаемость (т.е. реабсорбцию) стенки протоков для мочевины. Комбинации этих эффектов результируют осмоляльность вторичной (дефинитивной) мочи.
ПОЧКИ И КИСЛОТНО-ЩЕЛОЧНОЕ РАВНОВЕСИЕ
Лёгкие и почки имеют первостепенное значение для поддержания кислотно-щелочного равновесия крови путём контроля за компонентами её буферных систем - CO 2 и HCO 3 - . Кислотно-щелочное равновесие рассмотрено в главе 28, контроль - в главе 25, в этом разделе разобрана роль почек в контроле плазмы крови и в экскреции нелетучих кислот.
Нелетучие кислоты. В организме образуются нелетучие кислоты: (например, серная, фосфорная и различные органические) в суммарном количестве (за вычетом нейтрализованных основаниями) около 70 ммоль/сут (1 ммоль/кг массы тела). В перерасчёте на угольную кислоту (H 2 CO 3) почка ежесуточно экскретирует около 70 ммоль of H+ в мочу и одновременно переносит в кровь 70 ммоль вновь образованного HCO 3 - . В крови HCO 3 - нейтрализует 70 ммоль нелетучих кислот.
Титрование отфильтрованного HCO 3 - . Ежесуточно обе почки отфильтровывают 4320 ммоль HCO 3 - . Этот огромный пул анионов практически не экскретируется и не реабсорбируется, а титруется секретируемым в просвет канальцев H+ до CO 2 и H 2 O (H+ + HCO 3 -
H 2 CO 3 , H 2 CO 3 -- H 2 O + CO 2). Однако, реакция протекает слишком медленно для быстрого и полного превращения HCO 3 в H 2 O и CO 2 . Поэтому в процесс нейтрализации включается карбоангидраза эпителия почечных канальцев (фермент расщепляет HCO 3 - на CO 2 and OH - , а секретируемый H+ нейтрализует OH - , в итоге образуются те же H 2 O и CO 2 .).
«Новый» НСO 3 . Поверхностная мембрана эпителия хорошо проницаема для CO 2 и воды, поэтому CO 2 и H 2 O диффундируют в клетку, где карбоангидраза катализирует обратную реакцию - образование H + и HCO 3 - из CO 2 и H 2 O. Клетка экспортирует H + в просвет канальцев, а HCO 3 - в кровь через интерстиций. Таким образом, взамен оттитрованного в просвете канальцев и на поверхности эпителия HCO 3 - появляется «новый» HCO 3 - , секретируемый в кровь.
Титрование отфильтрованного и секретированного аммиака. Секретируемый H+ также титрует NH 3 . Небольшая часть NH 3 отфильтровывается, значительная часть диффундирует через эпителиальные клетки и поступает в просвет при помощи Na-H-обменника. В проксимальных канальцах превращение глутамина в α-кетоглутарат приводит к появлению 2 ионов NH 4 + , образующих 2 NH 3 2 иона H + . при метаболизировании α-кетоглутарата образуется 2 иона OH - , которые карбоангидраза превращает в ион HCO 3 - . Этот «новый» HCO 3 - далее поступает в кровь.
Титрование других отфильтрованных анионов. Помимо аммиака и HCO 3 - секретированный H + титрует также отфильтрованные HPO 4 - , креатинин и ураты.
Таким образом, в просвете канальцев H + титрует HCO 3 - , HPO 4 2- , NH 3 и некоторые другие анионы. Из 4390 ммоля H + 4320 ммоля (98%) идёт на титрование HCO 3 - . В итоге образуется «новый» HCO 3 - , поступающий в кровь. Эти процессы происходят преимущественно в проксимальном отделе нефрона (80%).
